������ѧѧԺ��Ӧ��������ʾ��������ϸ�����鵰��ȥ����ø���������Լ�����Ⱦɫ��Ũ���ķ��ӻ���
Ⱦɫ��Ũ����ϸ�����ѵ�ǰ���������˿���Ѻͼ������Ѷ���Ҫ�����ƺ��ϸ��Ⱦɫ��Ũ���ɰ�״��Ⱦɫ�壬������֤���ڵ���ȷ���롣���֮�£���˿������Ⱦɫ���ǰ�ڵ������ǿ��ٵ�һ��Ũ�������������Ѱ�����������I��II�����м�������I�Ǹ�ϸ�����ѷ�ʽ�����У�ǰ��I�ְ���ϸ���ڡ�ż���ڡ������ڡ�˫���ں��ձ��ڣ�������Ҫ�漰����ĸͬԴȾɫ��֮�����ԡ���������顣Ϊ�˱�֤��Щ�����¼���������У�Ⱦɫ���Ũ�������еġ��о�������Ⱦɫ��Ũ����Ҫ��Ũ����ճ𤵰��������ƣ����������������ѽ�������ε���Ũ���������壬�Ӷ����Ⱦɫ�����Ũ�������в��������Ӧ��������Ŷ�2016���ʾ��ֲ���������ϸ���������PHD�ṹ��MMD1�������ͨ��PHDʶ���鵰��H3K4me3ֱ�ӵ���Ũ���ǻ�CAP-D3�ı���Ӷ�������Ⱦɫ���ǰ��Ũ�����̣�Wang et al., Plant Cell����Ȼ����MMD1����CAP-D3�ľ�����ӻ����Բ������
6��22�գ�������ѧ������ѧѧԺ�о�Ա��Ӧ�������������������������������ѧ (Pennsylvania State University) ���������ڡ���Ȼ-ֲ���Nature Plants����������Ϊ��Cell-type-dependent histone demethylase specificity promotes meiotic chromosome condensation in Arabidopsis�����о����ģ���ʾ�˼�������ϸ�����鵰��ȥ����ø���������Լ�����Ⱦɫ��Ũ���ķ��ӻ��ơ�

Ϊ�˽���MMD1�鵼������������Ⱦɫ��Ũ�����̵ĵ��ػ��ƣ��о���Ա��2010�꿪ʼ������ͬ��MMD1�ض̵��ײ�ɸѡ�Ŀ⣬��2011�����������һ����������JMJ16�������һ��H3K4���鵰��ȥ����ø��ͼ 1�����о��Ŷ��������ר�ҡ��п�Ժ�Ŵ��뷢������ѧ�о���������Ժʿ������֤����JMJ16��������ֻ��ȥ��H3K4me2/3�������о�������������������鵰��������Ҫ�ٽ�����ı��ȥ�������ƻ���ı���⽫��������MMD1�������ٽ�CAP-D3����ı��

ͼ1 MMD1��JMJ16���Ľṹ�����������
�о���Ա�Ʋ⣬MMD1����Ӱ����JMJ16��ø���������ʵ���֤��MMD1ȷʵ������JMJ16�ĵ��������ԣ� MMD1-JMJ16���������ȥ���Ķκͺ�С���ϵ�H3K9me3����ôMMD1������ε�����JMJ16�ĵ����������أ��������������ʵIJ�ͬ�ض̵��ף�������MMD1��MMD�ṹ���Զ��壬ֲ���б��أ���JMJ16��C��FYR-C�ṹ������ͼ 1������ʾ��JMJ16��C�˿���Ӱ������ø��ṹ��Ϊ����֤������裬�о���Ա������ֻ����JMJ16-N�˴��ṹ��Ľض̵��ף�����JMJ16-N����ͬʱʶ���ȥ��H3K4me3��H3K9me3����һ������֤��JMJ16-N��JMJ16 ��C��FYR-C�ṹ������ã�����FRETʵ��Ҳ֧��JMJ16�����˿��ĺܽ���ֱ�ӻ�����ͼ2����
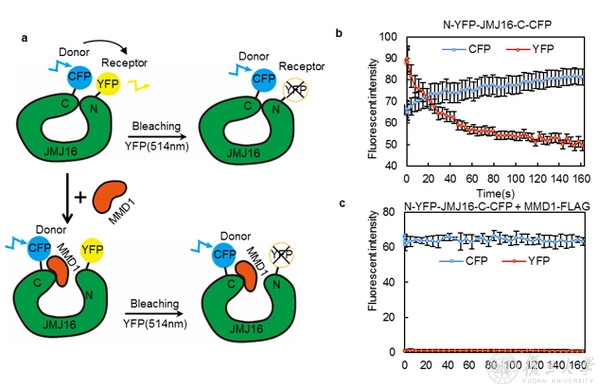
ͼ2 JMJ16-N��-C�˻�����MMD1���以����Ӱ��
��ôMMD1�����Ӱ��JMJ16�������˵Ļ����أ�ͨ�����⾺��ʵ�����MMD1���Ժ�JMJ16-N�����ԵĽ��FYR-C�ṹ�Ӷ����FYR-C����JMJ16-N������ЧӦ������FRET-bleachingʵ���ʾMMD1��������JMJ16��N�˺�C�˵ľ��룬������������� ��ͼ 2�������Ͻ��֤����JMJ16-C�˽ṹ�������������N�˴��ṹ��ʶ��H3K9me3����MMD1������JMJ16��C�˾�����ϣ�����ͨ��пָ�ṹ��C5HC2�����������ЧӦ ��ͼ 3�����ٽ�JMJ16ʶ���ȥ��H3K9me3��

ͼ3 ӫ��ƫ����� JMJ16ø��ṹ��пָ�ṹ���H3K4me3��H3K9me3�Ľ��ǿ��
jmj16�ļ�������ϸ��ת¼������֣�ҶƬ����Ҫ��������ع��ܻ���ı������������ϸ���б��ֺ�MMD1��ͬ�ٽ�һ������ı������Ⱦɫ��Ũ����ػ�����Ŀ�Ļ�����CAP-D3�������������鵰�����Σ�����H3K4me1/2/3 �仯����������H3K9me3�������ӡ����ڶ�����JMJ16��ͬԴ����KDM5/JARID��������PHD�ṹ���о���Ա�Ʋ�JMJ16�Ķ�λ��������MMD1��PHD�ṹ��ʶ��H3K4me3����MMD1��MMD�ṹ����ļJMJ16������Ӱ������������ԣ���������Ŀ�Ļ���ı��Ϊ����֤��һ���裬�о���Ա������JMJ16��ø��ṹ���ں�MMD1��PHD�ṹ��ת����mmd1��ͻ���壬���ֿ��Բ��ָֻ�mmd1Ⱦɫ��Ũ���쳣�ı��͡�
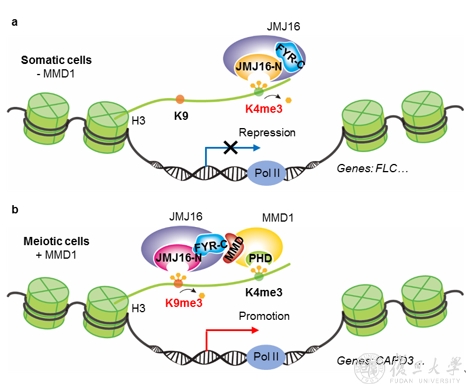
ͼ4 JMJ16��MMD1�ڲ�ͬϸ�������е��ػ�������ģ��
�����������ۣ��о���Ա�����JMJ16��MMD1����ϸ���ͼ�������ϸ���е��ػ�������ģ�͡�����ϸ���У�JMJ16��N�˺�C�˻�������������ʶ��H3K9me3��������ֻ����H3K4me3ȥ�������ԣ��Ӷ����ư���FLC���ڵĻ�����ͼ4 a�������ڼ�������ϸ���У�MMD1��PHDʶ��H3K4me3����MMD�ṹ����JMJ16��FYR-C�����������������C�˺�N�˵Ļ�������չ��JMJ16��H3K9me3ȥ�������ԣ��Ӷ��ٽ�����CAP-D3���ڵĻ�����ȷ����������Ⱦɫ�����ȷŨ����ͼ4 b�����ý��Ҳ�ǵ�һ��֤���˺���JmjC�ṹ���ȥ����øø����������Եĵ��ػ��ƣ���ʾ��ȥ����ø�ڲ�ͬ��ϸ�������п����벻ͬ��������Эͬ��ȷ������ػ���ı�������ֵ��ػ��ƿ��ܾ����ձ��ԡ�
��Ӧ�顢�����������DZ��о��Ĺ�ͬͨѶ���ߡ�������ѧ�ѱ�ҵ��ʿ���������೬��Ϊ���ĵĹ�ͬ��һ���ߡ��п�Ժ�Ŵ��뷢���о���������Ժʿ�����ʱ�ʿ���������˹���������Ŀ�õ�������Ȼ��ѧ�ܳ�������𡢹����ص�����о��ƻ���Ŀ������ѧ�Ŵ����������ص�ʵ���ҵĴ���֧�֡�
�������ӣ�https://doi.org/10.1038/s41477-020-0697-0
δ����������ת�أ���ѧ�Ż� » ������ѧѧԺ��Ӧ��������ʾ��������ϸ�����鵰��ȥ����ø���������Լ�����Ⱦɫ��Ũ���ķ��ӻ���
����Ƽ�
- ������ҽ�쵼���ӿ���ο��Ԯ�ظɲ�
- ���ҽ�ѧ��������ѧϰ�ؼ���¯
- ����Ų��������ߣ��������ſγ������µ��á�5��֮��
- ���������о�������������ѧ��ʮһ�조ѧ��֮�ǡ��صȽ�
- �������й�����70����ȫ�濪���¸�� ����츴����ϯ����ѧ����̳����
- ҧ��Ŀ���ٳ�����������ѧ��Զ����18��ɹ���2018����Ϻ��п�ѧ������
- ���������˿�Խ�����ֺ蹵������������ѧԺ����Ϣ���ǡ�ʵ���ӿ���������������ġ����ޡ�
- ������ѧ���ع������һ�λ�Ա��������ٿ�
- ������ѧ���гɹ������20���й����ʸ��¼����ɹ�����
- ���з��о�Ա�����顢����½��ڿ���������о� ������������ĤѪ�ܲ����о�����ȡ���½�չ
- ������ѧ����/��˼/����ܿ/����ΰ�Ŷӷ���Ԥ�Ⲭ��������Ե�����Һ�����־��
- ������ѧ��ǡ������ܡ�����ǿ�����Ŷ��о�����RNF43�ڽ�ֱ��ӡ��ϸ�����з�����Ƶͻ��
- ������ѧ���˴��������ЭίԱ��2021���������ϻ�����ְ
- ����һ���붬���������������еĸ���
- �������λ������� ��������������ʦ�ܺ�����ô����
- ��������ڼ临����ѧ��Уѧ����Ϊ�淶����
- ����ѧ�����Ͻ�����ѯ�����Ʒ���������������
- �Ϻ��С����С��ý�ʦ���������ڳ��о����Դ����̣�ڹ��ʱ������������
- ������ѧ����ίԱ������ְ������������
- ����ϵĸ��������㿴�鸴��ʳ����ʳ
���Ź���
- ������ҽ2022��2�¿��гɹ�һ�� 03-15
- ������ҽ������ʱ�裡 03-14
�߿�����
- ������ѧ2017�������³� 08-05
- ������ѧ2018�������³� 08-05
- ������ѧ2014�������³� 08-05
- ������ѧ2015�������³� 08-05
- ������ѧ2016�������³� 08-05
- ������ѧ2012�����³� 08-05
- ������ѧ2013�������³� 08-05
- ������ѧ2008�������³� 08-05
- ������ѧ2009�������³� 08-05
- ������ѧ2011�������³� 08-05